
Yohimbe
Tratto da “Piante Medicinali – Chimica, Farmacologia e Terapa” di R. Benigni, C. Capra e P.F.Cattorini
(Corynanthe Yohimbe (o Yohimba) K. Schum.
Fam. – Rubiacee/Cinchonee)
(Sin. – Pausinystalia Yohimba (K. Schum.) Pierre – Pseudocinchona Yohimbe K. Schum.)
Yohimbe- Ultimo aggiornamento pagina: 27/02/2018
Indice dei contenuti
- Generalità
- Componenti principali
- Proprietà farmacologiche
- Estratti e preparati vari
- Preparazioni usuali e Formule
- Bibliografia
Generalità

Etimologia – Corynanthe dal greco korunh = clava e anqos = fiore, per i boccioli fiorali che hanno forma di clava.
Yohimbé – nome indigeno della pianta.
Pausinystalia, dal greco pausis = pausa, fermata, cessazione e nustaxis = sonnolenza, da pauw = faccio cessare, allontano e nustazw = mi addormento, sonnecchio. Pianta sonnifuga, che vince la sonnolenza.
Pseudocinchona, dal greco yeudos = falso e cinchona = la china (corteccia di), cioè falsa china.
Nota – In realtà i generi Corynanthe, Pausinystalia, Pseudocinchona sono diversi l’uno dall’altro, per quanto vicinissimi. Il gen. Pseudocinchona non è riconosciuto da alcuni botanici.
Nomi volgari indigeni – Yohimbé, yohimba.
Habitat – Gabon, Camerun. L’indicazione Congo è errata. Nel Congo esistono altre specie del genere, ma non la Yohimbé.
Grande albero della foresta equatoriale.
Parti usate – La corteccia.
Componenti principali
Gli alcaloidi della corteccia del Corynanthe Yohimbe sono basi terziarie indoliche, distinguibili in due gruppi:
1) gruppo della yoimbina e dei suoi isomeri (corinantina, pseudo-yoimbina, allo-yoimbina, a-yoimbina e b-yoimbina);
2) gruppo della corinanteina e della tetraidroalstonina (ajmalicina, corinanteina e diidrocorinanteina).
1) Gruppo della yoimbina e isomeri
Yoimbina (quebrachina), C21H26O3N2, estratta per la prima volta da Aspidosperma quebracho-blanco e denominata quebrachina da Messe (1) nel 1882, è stata ottenuta da Spiegel (2) nel 1890 dal Corynanthe Yohimbe (Pausinystalia Yohimba). Più recentemente è stata ricavata da diverse specie di Rauwolfia (ved. tabella 1; ved. anche Rauwolfia serpentina).
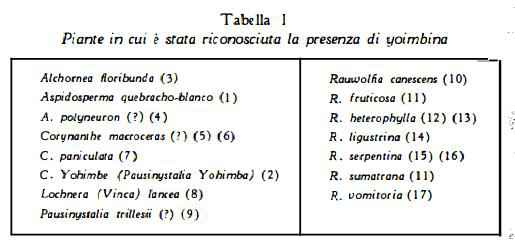
L’identità della quebrachina e della yoimbina fu a lungo discussa (18- 20), ma è stata poi definitivamente dimostrata (21-24).
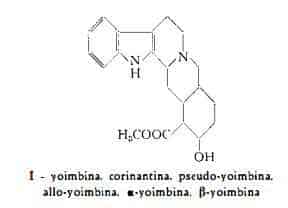
La formula grezza, C22H30O4N3, proposta da Spiegel (2), è stata successivamente corretta nella C21H26O3N2 (25) (26). Oggi pienamente nota, la formula di struttura I della yoimbina fu oggetto dello studio di numerosi AA. (27-58) per oltre vent’anni. Alla sua definizione hanno particolarmente contribuito i risultati delle ricerche sulla composizione della yobirina, della tetraidroisoyobirina e della chetoyobirina.
Corinantina (rauimbina) era già stata estratta nel 1909 da Fourneau (59) dalla Pseudocinchona africana, quando fu trovata anche nella Corynanthe Yohimbe da Jorio (60) nel 1939. E’ stata identificata da Hofmann (16) con la rauimbina isolata dallo stesso A. (1) dalla Rauwolfia serpentina Benth. E’ uno stereoisomero levogiro della yoimbina (62) (63), la cui formula è stata confermata per sintesi (64).
Pseudo-yoimbina (yoimbene) è lo stereoisomero della yoimbina (62 ), che fu denominato yoimbene, quando fu isolato per la prima volta dalle acque madri, ottenute dalla preparazione industriale della yoimbina (65) (66). Con il nome di pseudoyoimbina è stato designato più recentemente, quale componente della Rauwolfia canescens (67) e della Rauwolfia tetraphylla (68). Janot, Goutarel e Amin (69) hanno determinato l’identità dello yoimbene e della pseudoyoimbina. La differenza tra le azioni farmacologiche delle due sostanze, segnalata da Raymond-Hamet (70), è stata riferita da Janot e coll. (62) all’azione di altre sostanze, presenti come impurezze nei campioni esaminati.
Allo-yoimbina, altro stereoisomero della yoimbina (71), è stata separata dalla corteccia del Corynanthe Yohimbe da Warnat (72) e da Hahn e Brandenberg (73). La sua composizione, quale stereoisomero della yoimbina è stata confermata dall’identificazione della yobirina, tetraidroisoyobirina e della chetoyobirina nei prodotti di deidrogenazione con selenio (71).
a-Yoimbina (corinantidina, rauvolscina) è lo stereoisomero della yoimbina (74), ottenuto per la prima volta dal Corynanthe Yohimbe e denominato perciò corinantidina (18) (72) (75) (76). Ha assunto anche il nome di rauvolscina, come componente principale della Rauwolfia canescens (77), finché l’identità della rauvolscina e della a-yoimbina è stata stabilita in seguito allo studio dei prodotti di degradazione delle due sostanze (78-85). L’a-yoimbina è stata ottenuta anche dalla Alstonia constricta (86), Pseudocinchona africana (87), Rauwolfia heterophylla
(13), Rauwolfia hirsuta (canescens) (88), Rauwolfia ligustrina (14), Rauwolfia sumatrana (11). Rauwolfia vomitoria (17). Serpina fu denominata una sostanza che è stata estratta dalla Rauwolfia serpentina (89) (90) e dalla R. canescens (91) e che Hochstein (92) ha dimostrato essere costituita da «cristalli misti» di yoimbina e di a-yoimbina.
b-Yoimbina (amsonina), stereoisomero della yoimbina (93) (94), è stata ottenuta da Heinemann (95) dalla corteccia del Corynanthe Yohimbe e più recentemente da Hofmann (96) dalla Rauwolfia canescens. Il nome di amsonina è stato dato ad essa da Kimoto e coll. (97) (98), che l’hanno isolata dalla Amsonia elliptica.
2) Gruppo della corinanteina e della tetraidroalstonina
Ajmalicina, C21H24O3N2, è stata in un primo tempo isolata dalla Rauwolfia serpentina Benth. da Siddiqui e Siddiqui (99) nel 1931 ed in seguito da numerose altre specie di Rauwolfia. Con il nome di d-yoimbina è stata designata nel 1934 da Heinemann (95), che l’ha estratta dalla corteccia dello Yohimbe.
L’identità dell’ajmalicina e della d-yoimbina è stata stabilita da Neuss e coll. (100) e da Janot e Le Men (101). La formula di struttura (II)
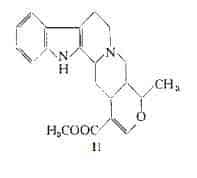
della ajmalicina fu definita da Goutarel e Le Hir (102) nel 1951 ed è stata poi confermata per sintesi in seguito agli studi volti alla determinazione della configurazione spaziale (103-105) ed alla sintesi del prodotto racemico (106).
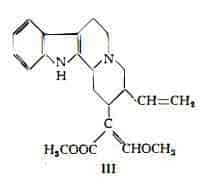
Corinanteina, C22H26O3N2, è stata estratta dapprima dalla Pseudocinchona africana (59) (107) e poi dal Corynanthe Yohimbe (108). Dallo studio dei prodotti di degradazione e di trasformazione della corinanteina (109-113) si è dedotta la formula di struttura III (112-120).
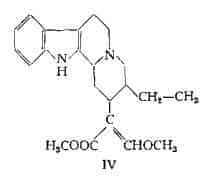
Diidrocorinanteina, C22H28O3N2, fu separata da Karrer, Schwyzer e Flam (114) dalla corinanteina, estratta dalla Pausinystalia Yohimba e dalla Pseudocinchona africana. La diidrocorinanteina ha la formula strutturale IV, confermata dalla sintesi totale del prodotto racemico (122).
Contenuto di alcaloidi
La quantità percentuale di alcaloidi nelle cortecce commerciali oscilla entro limiti piuttosto ampi. Sono stati segnalati titoli di alcaloidi totali, varianti dallo 0,5 al 3,2% fino al 6%, secondo Brandt (122), ed anche dal 5,35 al 9,35 % in cortecce originarie del Cameroun, secondo le analisi di Perrot e Raymond-Hamet (123).
Per il titolo di yoimbina della corteccia dei fusti sono stati ottenuti valori pari a 1-1,5% (124), anche 0,36-1,195% (125), 0,480-0,674% (126) e, per cortecce provenienti dal Cameroun, 1,67-3,4% (123).
Le differenze, riscontrate nel titolo alcaloidico delle cortecce commerciali, sono state anche messe in relazione con il diverso rendimento che si ha a seconda del procedimento usato per l’estrazione (7). Variazioni sensibili sono state inoltre notate per il titolo della corteccia in rapporto alla posizione che questa occupa sull’albero, Le rese migliori sono state date dalla corteccia dei fusti più sottili e dalla corteccia delle radici, dalla quale è stato ricavato circa il 5 % di alcaloidi (127). Yoimbina ed i suoi isomeri sono stati trovati in quantità crescenti dalla base alla cima del tronco; i rami e le foglie ne contengono piccole quantità (128). Nella corteccia del fusto il contenuto di alcaloidi è basso verso la base dell’albero, aumenta progressivamente fino ad un massimo alle prime diramazioni, quindi diminuisce gradatamente lungo il ramo fino alle foglie, nelle quali gli alcaloidi sono in piccole quantità e differiscono qualitativamente da quelli della radice e del fusto. Le foglie, oltre alla yoimbina, che in esse non è più l’alcaloide principale, contengono numerosi derivati indolici, che forse rappresentano stadi della sintesi della yoimbina (127).
Proprietà farmacologiche ed impiego terapeutico
Presso alcune popolazioni del Congo la corteccia di Yohimbe è usata come stimolante nervoso, per resistere al sonno e, analogamente all’uso che fin dall’epoca precolombiana, gli indigeni delle regioni andine del Nord dell’Argentina, fanno della corteccia di Quebracho blanco, la corteccia di yohimbe è impiegata dai nativi dell’Africa centrale come antimalarico. Presso questi ultimi inoltre, la droga gode fama di afrodisiaco, attività che venne in parte confermata per il suo alcaloide principale, la yoimbina, dalla successiva sperimentazione clinica e farmacologica [Observate e Loewy (129), Muller (130)] e venne attribuita ad un’eccitazione dei centri lombari (Muller) o a variazioni circolatorie locali [Raymond-Hamet (131)], ma non è dimostrato che questo effetto possa essere posto in relazione con una eventuale azione endocrina dell’alcaloide. Non sembra comunque che la produzione degli spermatozoi venga stimolata e che il suddetto effetto possa essere prodotto, almeno nell’uomo, con dosi terapeutiche.
Dosi di 1-2 mg provocano nelle ratte un prolungamento dell’estro e dosi più elevate (2-4 mg) determinano la comparsa di una pseudogravidanza, effetti questi che sono stati spiegati da Fugo e Cross (132), con una stimolazione dell’ipofisi. Il Ritmo di riproduzione nei topi, non viene però aumentato [Rumry (133)].
Azione adrenolitica
Questa azione della yoimbina è forse la più interessante dal punto di vista farmacologico e la sua scoperta viene di solito attribuita a Raymond- Hamet il quale nel 1925, comunicò all’Academie des Sciences di Parigi le sue osservazioni «su di un nuovo caso d’inversione degli effetti adrenalinici» (134), concludendo che la yoimbina, contrariamente ai principi fisiologicamente attivi della Segale cornuta, è ben definita, poco costosa, poco tossica, dà luogo a sali stabili, ha un’azione durevole e particolarmente adeguata allo studio degli adrenalinici e della dissociazione del simpatico motore dal simpatico inibitore. L’esistenza di vasodilatatori simpaticolitici essendo ben dimostrata farmacodinamicamente, si deve ammettere, secondo l’A., che oltre l’antagonismo simpatico-parasimpatico, esista anche un antagonismo simpatico motore-simpatico inibitore che sembra essere sfuggito a molti clinici. In realtà, sette anni prima Filippi (135) studiando comparativamente l’azione della quebrachina e della yoimbina, concludeva che questa non poteva essere identificata con quella, distinguendosi la yoimbina per la sua più forte azione inibitrice dell’ipertensione adrenalinica, per l’azione paralizzante che essa possiede sul simpatico cervicale, azione di cui non sarebbe dotata la quebrachina ecc. (vedi Quebracho blanco) il che dimostra che l’azione adrenolitica della yoimbina era già nota, anche se le conclusioni cui giunse Filippi sulla non identità delle due sostanze, vennero poi confutate successivamente dallo stesso Raymond-Hamet (136-137) il quale giunse invece alla conclusione che le due sostanze sono identiche sia per la loro costituzione chimica che per la loro attività farmacologica.
La proprietà della yoimbina d’invertire l’azione di determinate dosi di adrenalina e la sua azione vasodilatatrice, venne successivamente studiata nei suoi particolari dallo stesso Raymond-Hamet (138-144) e confermata da altri autori, quali Hutchinson e coll. (145), Barry e Chauchard (146), Barry (147), Koppanyi e coll. (148) ed altri.
Questa attività simpaticotropa della yoimbina si riflette naturalmente sulle funzioni cardiovasale, respiratoria e di altri settori del sistema nervoso autonomo.
Per quanto riguarda l’attività sul sistema cardiovascolare e respiratorio, Raymond-Hamet (138), dimostrò, sperimentando sui vasi renali, che la yoimbina paralizza il simpatico motore (vasocostrittore) senza modificare l’eccitabilità del simpatico inibitore (vasodilatatore).
Egli trovò, che i vasocostrittori renali hanno una sensibilità particolare, non solo all’azione dell’adrenalina [Jonescu (149)], ma anche a certi antagonisti di essa, quali la yoimbina, gli alcaloidi della Segale e l’atropina (140).
Lo stesso Raymond-Hamet, l’anno seguente, studiò comparativamente l’inversione degli effetti adrenalinici prodotta dalla yoimbina e dagli alcaloidi della Segale cornuta (139). Contrariamente a questi ultimi che sono vasocostrittori, la yombina è vasodilatatrice.
Nel cane cloralosato inoltre, con la yoimbina si ottiene sempre l’inversione dell’ipertensione prodotta da piccole dosi di adrenalina (5/1000 di mg) mentre con l’ergotamina è spesso impossibile ottenerla.
Per contro, sempre nel cane cloralosato, l’inversione ottenuta con ergotamina è tanto più accentuata quanto più la dose di adrenalina è elevata, mentre con la yoimbina è difficile invertire completamente l’azione di dosi elevate di adrenalina (da 0,5 a 1 mg). Infine, contrariamente agli alcaloidi attivi della Segale cornuta i quali, secondo Dale (150), permettono d’abolire l’azione cardioacceleratrice dell’adrenalina, la yoimbina non influisce su questa reazione.
Nel cane cloralosato, sottoposto all’azione della yoimbina, l’adrenalina sebbene conservi la sua azione vascolare normale, determina nella maggior parte dei casi, un’ipotensione immediata e non preceduta, come generalmente avviene con gli alcaloidi della Segale, da un’ipertensione transitoria. E’ quindi difficile, secondo l’A. ammettere con Dale (151), che questo effetto ipotensivo transitorio prodotto molto frequentemente dall’adrenalina nell’animale ergotaminizzato, sia dovuto in questi casi, al suo potere cardioacceleratore.
Contemporaneamente Houssay e Molinelli (152) trovarono che la yoimbina inverte l’effetto ipertensivo della nicotina anche in assenza delle surrenali e che aumenta l’effetto ipertensivo dell’eccitazione dello splancnico.
L’azione adrenalinosecretrice che consegue all’eccitazione dello splancnico e all’iniezione endovenosa di nicotina, viene ugualmente inibita dalla yoimbina, dalla quale però, la secrezione adrenalinica non è eccitata.
Tanto la yoimbina quanto l’adrenalina, somministrate a dosi opportune nel cane cloralosato, inducono, secondo Gley e Czarnecki (153), apnea; ma se dopo la somministrazione di yoimbina, si somministra adrenalina a dosi sicuramente apneiche, quando la registrazione indica il ritorno alla attività normale, non si ha più apnea ma polipnea. Altrettanto avviene se l’apnea è provocata da adrenalina.
L’apnea prodotta da nicotina non è modificata dalla yoimbina, ma è impedita e invertita dall’ergotamina, così come l’apnea prodotta dall’ergotamina è ostacolata e invertita dalla nicotina.
Loeper e Lemaire (154) hanno osservato che l’acetilcolina somministrata per via endovenosa, produce sulla pressione carotidea del cane, un effetto depressivo più duraturo, dopo yoimbinizzazione. Per dosi non superiori ai 0,33-0,50 mg, la durata dell’effetto ipotensivo passa da 5’ nell’animale normale, a 20’ e più, dopo somministrazione preventiva di yoimbina alla dose di 2 mg/kg.
Lo stesso effetto si ottiene quando l’acetilcolina e la yoimbina vengono iniettate contemporaneamente alle dosi di 2,5 e di 5 mg rispettivamente. In questo caso la durata dell’effetto ipotensivo è inferiore, ma tende a aumentare a mano a mano che le iniezioni vengono ripetute.
Un analogo prolungamento dall’azione ipotensiva si ottiene quando l’acetilcolina è iniettata per via sottocutanea nel cane yoimbinizzato, a dosi che non superino 1-2 mg/kg. In questo caso però compaiono sulla curva emodinamica, grandi onde d’ipertensione relativa. Nello stesso tempo si eleva anche la pressione endorachidea con onde che si intercalano con quelle della pressione ematica, disegnando una curva che rappresenta in qualche modo, l’immagine speculare della pressione carotidea.
Loeper e coll. (155) impiegando derivati della colina, hanno poi confermato i risultati ottenuti da Raymond-Hamet (142) nel cane atropinizzato. In queste condizioni l’acetilcolina determina un’ipertensione che secondo gli AA.. sarebbe dovuta ad una adrenalinemia secondaria e che per ciò può essere invertita dalla yoimbina.
Nel cane decapsulato, la somministrazione di yoimbina non aumenta gli effetti ipotensori dei derivati dell’acetilcolina. Essi hanno inoltre notato, che nel cane atropinizzato e yoimbinizzato, l’iniezione endovenosa di metilacetilcolina, determina una caduta della pressione. Heymans e Regners (156) dimostrarono che l’ergotamina sopprime i riflessi vasomotori del seno carotideo ed esagera nel cane, gli effetti cardioinibitori di questo riflesso, ciò che venne più tardi confermato da Nickerson (157).
Successivamente Heymans e Bouckaert (158), in considerazione delle analogie farmacologiche esistenti fra ergotamina e yoimbina, hanno voluto indagare se anche quest’ultima è capace d’influenzare nella stessa maniera, i riflessi vasomotori.
Essi hanno dimostrato infatti che la yoimbina sopprime questi riflessi e inibisce la reazione vasodilatatrice consecutiva all’eccitazione elettrica del nervo carotideo. I riflessi respiratori, al contrario di quelli vasomotori, non vengono invece influenzati.
Analogamente all’ergotamina, la yoimbina inibisce l’ipertensione asfittica ma non quella prodotta da pituitrina o da b-tetraidronaftilamina [Bouckaert e Czarnecki (159)].
Raymond-Hamet (143) ha inoltre dimostrato che una dose di adrenalina capace di produrre una molto netta vasocostrizione nell’animale normale, provoca al contrario, una vasodilatazione nell’animale pretrattato con yoimbina, confermando così che nei mammiferi l’azione vasocostrittrice dell’adrenalina è trasformata dalla yoimbina in azione vasodilatatrice, ciò che venne in seguito confermato da Barry e Chauchard (146) i quali trovarono che la yoimbina antagonizza l’effetto dell’adrenalina sulle arterie.
I vasi coronari, secondo Rabe (160) non reagiscono all’azione della yoimbina mentre, secondo Meyer (161), essi verrebbero dilatati.
Somministrata per via orale o ipodermica, la yoimbina, a dosi moderate, determina secondo Weger (162), una vasodilatazione generale dei vasi cutanei, delle mucose e, in particolare, degli organi genitali, fatto quest’ultimo cui, come già detto, viene attribuito l’effetto afrodisiaco.
Raymond-Hamet (144) infine, ha studiato valendosi di una tecnica personale, il grado proporzionale dell’attività simpaticolitica di otto isomeri della yoimbina, trovando che se per molti di essi tale attività è presso a poco uguale a quella della yoimbina, essa è 17 volte più debole per la corinantina è 131 volte più debole per la pseudo-yoimbina, ciò che dimostra che queste due sostanze sono inutilizzabili nella terapia delle simpatosi.
Le risposte alla yoimbina degli animali in anestesia sono diverse da quelle che si ottengono negli animali normali. Negli animali in anestesia la yoimbina determina una caduta della pressione che nei cani può raggiungere i 35 mmHg, mentre nelle pecore è notevolmente inferiore (10 mmHg).
La frequenza cardiaca nei cani in anestesia, subisce un aumento medio di 30 battiti/min., inferiore quindi a quello prodotto negli animali normali e non subisce variazioni significative dopo adrenalina o noradrenalina [Gershon e Lang (163)]. Nei cani anestetizzati la yoimbina inverte, come già detto, la risposta pressoria all’adrenalina mentre la risposta alla noradrenalina è invertita in alcuni casi e inibita in altri (164, 165, 166).
Gli animali non anestetizzati, reagiscono alla yoimbina in maniera diversa: nei cani e nelle pecore alla dose di 0,5 mg/kg, essa non determina una caduta della pressione come negli animali in anestesia, ma un aumento medio che nel cane raggiunge i 60 mmHg e 20 mm nella pecora.
La frequenza cardiaca aumenta come negli animali anestetizzati ma in misura maggiore, 80-90 battiti/min, nei cani [Gershon e Lang (163)] e ciò concorda con quanto è stato notato nell’uomo dallo stesso Gershon e da Holmberg (167).
Nei cani non anestetizzati la yoimbina induce modificazioni ECgrafiche che si manifestano con un aumento del potenziale dell’onda T e con tachicardia.
Secondo Gershon e Lang (163) queste variazioni presentano analogie con quelle che si ottengono sotto sforzo, se si ammette che esse siano dovute alla mobilizzazione delle catecolamine e trovano significativo il fatto che la sola adrenalina provoca tachicardia senza notevole aumento dell’onda T, mentre la noradrenalina, al contrario, determina un aumento molto maggiore dell’onda T e meno tachicardia.
La frequenza cardiaca in risposta all’adrenalina o alla noradrenalina, dopo yoimbina, sembra essere dovuta, secondo Gershon e Lang, a un riflesso dipendente dalle variazioni della pressione.
L’inversione della risposta all’adrenalina è stata dimostrata anche nei cani non anestetizzati, ma in questo caso la risposta pressoria alla noradrenalina veniva soltanto inibita ma non invertita.
Nei cani, nelle pecore e nei gatti non anestetizzati, la yoimbina produce un apparente prolungamento dell’anestesia con risposte motorie e un miglioramento della frequenza e del volume respiratorio.
Shaw e Wolley (168) hanno trovato che la yoimbina, analogamente ad altri derivati indolici strutturalmente simili alla serotonina, antagonizzano l’azione vasocostrittrice di quest’ultima e ne invertono l’azione. Nei cani e nelle pecore non anestetizzate, essa produce una diminuzione della risposta alla serotonina e, soprattutto, una minor durata di essa, con rapido ritorno alle condizioni normali (163).
Northover e Subramanian (169), i quali hanno esaminato un gruppo di sostanze in relazione alla loro capacità di antagonizzare l’edema indotto dalla formaldeide o dalla serotonina nella zampa del topo, hanno trovato, per quanto riguarda la yoimbina, che questa, alla dose di 4 mg/kg, riduce del 50% la reazione edematosa.
Azione psichica – Come altri alcaloidi indolici, anche la yoimbina può produrre alterazioni psichiche che sotto certi aspetti ricordano quelli prodotti da altri alcaloidi o sostanze strutturalmente simili (vedi Segale cornuta e Tabernanthe iboga).
Ricerche cliniche eseguite da Holmberg e Gershon (167), hanno stabilito che gli effetti autonomi prodotti dalla yoimbina, sono di natura adrenergica e che contemporaneamente a questi effetti autonomi, si possono osservare alterazioni psichiche che simulano uno stato ansioso, durante il quale i soggetti presentano una maggior irritabilità e, inoltre, che il grado di queste risposte è in relazione, secondo gli AA., col livello basale dell’attività emotiva.
In taluni soggetti schizofrenici la yoimbina può attivare un processo psicotico più o meno latente. Essa produce un aumento della reattività generale, delle allucinazioni attive, dell’espressione di delusione e delle manifestazioni dei disturbi del comportamento. Tuttavia, secondo gli AA., la yoimbina non è psicogena nei soggetti normali.
Nell’uomo le variazioni del comportamento sono correlate con lo stato ansioso, mentre nel cane esse sono in relazione alle risposte fisiologiche e si manifestano con tremori diffusi, aumento dello stato ansioso, ipersalivazione, diarrea, iperventilazione e dilatazione dei vasi congiuntivali.
Nelle pecore questa sintomatologia è meno evidente: esse divengono più vigilanti, con le orecchie ritte e le gambe tese (163, 167).
La risposta di tipo ansioso alla yoimbina è molto maggiore, secondo Gershon e Lang (163), di quella prodotta dall’adrenalina ed è di origine centrale. Gli stimolanti e i deprimenti centrali studiati dagli AA., inibiscono o potenziano rispettivamente, gli effetti autonomi e psichici della yoimbina, il che è interpretato dagli AA. come un’ulteriore prova che gli effetti autonomi e psichici da essa prodotti, sono per la maggior parte, mediati centralmente.
Il potenziamento della risposta ansiosa, sia comportamentale che psichica, è molto marcato quando l’azione della yoimbina viene sovrapposta a quella dell’imiprazina, le cui azioni centrali sarebbero, secondo Sigg (170), di natura adrenergica.
Secondo Gershon e Lang (163), la yoimbina può produrre nei cani uno stato ansioso modello che può essere considerato come Test per lo studio delle sostanze antiansia. Successivamente Holmberg, Gershon e Beck (171), sperimentando sull’uomo, hanno trovato che la yoimbina alla dose di 0,5 mg/kg endovena, corrisponde a quella massima generalmente ben tollerata.
Questa dose è ritenuta dagli AA. piuttosto bassa: essa non provoca alcun blocco adrenergico, ma solo alcuni effetti stimolanti centrali di grado variabile in relazione alla reattività emotiva individuale.
Gli AA., che hanno sperimentato su 15 pazienti schizofrenici e su 15 soggetti normali, hanno trovato che la yoimbina, somministrata a dosi opportune può, analogamente all’adrenalina, servire come Test per riprodurre stati ansiosi.
Azione sulla milza – Bariety e Kohler (172) hanno studiato comparativamente l’azione di diversi simpaticolitici sulla milza e, per quanto riguarda la yoimbina, essi hanno trovato che la sua azione può variare secondo la sensibilità individuale. Dosi di 3-4 mg/kg nel cane anestetizzato con cloralosene, possono determinare o una splenocontrazione notevole e transitoria oppure possono diminuire, aumentare leggermente o non modificare, il volume della milza.
Dosi superiori ai 3-4 mg/kg producono spesso splenodilatazione transitoria che sembra essere proporzionale alla dose somministrata. A volte si può osservare anche la comparsa di fatti convulsivi. Queste diverse reazioni si possono ottenere sullo stesso animale partendo da piccole dosi ed aumentandole progressivamente, purché fra un’iniezione e l’altra venga lasciato un intervallo di 10-20′.
Secondo gli AA., a queste azioni della yoimbina partecipano non solo le fibre muscolari liscie della capsula e delle travate spleniche, per l’azione che su di esse hanno le piccole e le forti dosi di yoimbina [Loewy e Rosenberg (173), Raymond-Hamet (174)], ma anche una componente vascolare dovuta all’elevata sensibilità dei vasi spleniti all’azione della yoimbina stessa [Raymond-Hamet (142)] e al fatto che piccole dosi dell’alcaloide (1 mg in toto), possono determinare un sia pur piccolo aumento del volume della milza, aumento che da A. e B. Chauchard e P. Chauchard (175), venne attribuito ad un fenomeno vascolare.
Azione sulla composizione del sangue
a) Sull’iperpotassiemia adrenalinica e noradrenalinica Delga e Hazard
(176) hanno trovato che la yoimbina oltre a dissociare gli effetti vasomotori dell’adrenalina sino ad invertirli, essa diminuisce leggermente anche il suo effetto sulla potassiemia, il cui aumento viene pertanto contenuto entro certi limiti.
Tuttavia a dosi convenienti, la yoimbina può dissociare gli effetti vasomotori dell’adrenalina sensa influire considerevolmente sulla potassiemia. A dosi sufficientemente elevate invece, la dissociazione non si manifesta più perchè la yoimbina può molto facilmente annullare o invertire leggermente l’azione della noradrenalina sulla potassiemia. L’effetto della yoimbina nel ridurre l’iperpotassiemia noradrenalinica, è dunque nettamente meno accentuato di quello da essa prodotto sull’iperpotassiemia adrenalinica (177).
Gli stessi autori (166), hanno studiato l’effetto della perfusione lenta con adrenalina e noradrenalina sulla pressione arteriosa, sulla potassiemia e sulla glicemia dei cani normali trattati con cloropromazina e con yoimbina. Dalle loro indagini risulta che nel cane la noradrenalina è più ipertensiva dell’adrenalina ma meno iperpotassiemizzante. L’azione dell’una e dell’altra sulla pressione arteriosa è indipendente dai loro affetti sulla potassiemia e sulla glicemia, i quali possono divergere perchè il loro meccanismo è diverso. Gli adrenolitici (cloropromazina e yoimbina) accentuano ancora questa dissociazione.
Per quanto riguarda la yoimbina essa, alla dose di 0,5-3 mg/kg, determina, all’inizio della perfusione con adrenalina o noradrenalina, la comparsa di un effetto ipotensivo la cui durata varia proporzionalmente alla rapidità della perfusione e alla dose di yoimbina.
Nel caso della noradrenalina, se la quantità dell’adrenolitico è debole (da 0,6 a 1 mg/kg), l’ipotensione è poco marcata e fugace; la pressione si rialza sotto l’apporto costante di noradrenalina (4 g/kg/min.) e si eleva a volte al di sopra della norma. Se la dose di yoimbina è elevata (3 mg/kg), l’ipotensione persiste per tutta la durata della perfusione.
Nelle condizioni in cui gli AA. hanno sperimentato, non sembra che la yoimbina abbia qualche influenza sull’iperglicemia adrenalinica o noradrenalinica. Quanto all’iperpotassiemia normalmente indotta da queste catecolamine, essa è considerevolmente attenuata o anche annullata dopo l’azione della yoimbina. Tuttavia, anche in questo caso gli AA. hanno constatato che l’azione iperpotassiemizzante dell’adrenalina, è più resistente di quella ipertensiva, all’azione della yoimbina.
Essi hanno potuto osservare infatti, il verificarsi di un’ipertensione e di una debole e contemporanea iperpotassiemia.
Cormier e coll. (178) hanno studiato l’azione della yoimbina sulle variazioni della composizione chimica del sangue nell’animale in ipotermia profonda.
Lo stato di ipotermia induce tre modificazioni plasmatiche molto caratteristiche e cioè, l’aumento considerevole e costante del glucosio, del potassio e del fosforo.
Lo stress da freddo libera un’importante quantità di adrenalina dalla midollare surrenale, la quale provoca a sua volta una notevole iperpotassiemia. Fenn e Asano (179), avevano già dimostrato che tutto il K liberato, non può provenire soltanto dalle perturbazioni del metabolismo glucidico.
Era quindi necessario stabilire, secondo Cornier e coll., se il K è liberato dall’adrenalina, indipendentemente dalla quantità relativa alle variazioni del metabolismo glucidico, come venne dimostrato da Silva (180) o se, in considerazione del fatto che l’azione del freddo non si limita soltanto a produrre un
’ipersecrezione di adrenalina, non è dovuto invece a qualche altro fattore.
Gli AA. in base ai risultati precedentemente riferiti, ottenuti da Delga e Hazard, i quali dimostrano che i simpaticolitici possono dissociare gli effetti adrenalinici, hanno prodotto un’ipotermia, in animali preventivamente yoimbinizzati e in queste condizioni essi hanno osservato che la potassiemia non subisce più un aumento, che la glicemia è molto debolmente ridotta e, inoltre, che la yoimbina non modifica la natriemia, la calcemia e la protidemia nell’animale in ipotermia profonda. Sembra dunque, secondo gli AA., che l’iperglicemia e l’iperpotassiemia prodotte dall’ipotermia, siano dovute allo stesso ormone, il quale agirebbe però, nei due casi, con un meccanismo d’azione diverso e i suoi effetti possono essere dissociati dai simpaticolitici e in particolare, dalla yoimbina.
b) L’azione della yoimbina sulla glicemia è stata studiata da Nitzescu e anche più estesamente, da Hanson.
Nitzescu (181) osservò che la yoimbina, come l’ergotamina, esercita un’azione inibitrice sull’iperglicemia simpatica conseguente a somministrazione di adrenalina. Questa azione sembra essere evidente soprattutto nel coniglio nel quale può essere completamente soppressa. Le piccole dosi di 0,5-1 mg/Kg di yoimbina, sufficienti per invertire l’effetto adrenalinico nel cane, non sembrano avere alcuna influenza inibitrice sull’iperglicemia adrenalinica. L’A. ammette che occorrono dosi diverse di yoimbina per esercitare un’azione paralizzante, secondo i diversi gruppi delle terminazioni del sistema nervoso simpatico.
Secondo Hanson (182), la yoimbina deve essere considerata come un agente debolmente iperglicemizzante. L’iperglicemia da essa prodotta è costante ma debole e di breve durata, in tutti i casi, essa scompare 3 ore dopo l’iniezione ed è seguita da ipoglicemia. Il ritorno alla norma avviene dopo la 3° o la 4° ora. L’A. spiega questa azione debolmente iperglicemizzante della yoimbina, ammettendo che essa, pur paralizzando il simpatico motore, possa invece eccitarlo a debole concentrazione, con conseguente stimolazione della glicogenolisi, e successiva inibizione; oppure che la yoimbina, per la sua azione sui muscoli scheletrici e stimolante del metabolismo muscolare, determini una mobilizzazione dei glucidi.
Successivamente lo stesso Hanson (183), ha studiato l’azione della yoimbina relativamente agli effetti dell’adrenalina sulla glicemia, dimostrando che essa può inibire completamente la mobilizzazione dei glucidi indotta dall’adrenalina e, inoltre, che essa può potenziare l’azione ipoglicemizzante dell’insulina (184).
Azione sui muscoli lisci – Fin dal 1914 Loewy e Rosenberg (173) notarono che l’intestino isolato o in situ e la vescica, sono stimolati da piccole dosi di yoimbina e depressi dalle forti dosi, ciò che venne confermato in seguito da Raymond-Hamet (174).
Più recentemente Thouvenat e Harichaux (185), hanno notato che quando un’ansa di digiuno-ileo di cavia è eccitata in due tempi con stimolazioni successive subliminari, è possibile seguire le modificazioni farmacologiche delle due fasi preparatorie e d’evacuazione del peristaltismo intestinale. L’atropina e l’esametonio, a deboli concentrazioni, aumentano il livello critico del tono longitudinale a partire dal quale compare l’attività ritmica.
Il rilasciamento della muscolatura longitudinale della zona orale, fra ogni contrazione ritmica, è relativamente ridotta in presenza di yoimbina e la trasmissione delle eccitazioni alla zona aborale è bloccata. Ciò, secondo gli AA., fa pensare all’intervento di neuroni adrenergici nella partecipazione della muscolatura longitudinale all’attività peristaltica nella sua fase ritmica.
Sulla muscolatura liscia dell’iride la yoimbina agisce determinando un effetto midriatico, senza perdita dell’accomodamento [vedi Sollmann (173)].
Azione antidiuretica – La yoimbina somministrata a cani normali esplica, secondo Fugo (186), un effetto antidiuretico che l’A. ritiene dovuto ad una scarica di ormone antidiuretico postipofisario. Infatti, allorché l’alcaloide viene somministrato a cani in cui il lobo posteriore dell’ipofisi è stato sezionato o asportato, l’azione antidiuretica manca.
Azione antipiretica – La yoimbina a piccole dosi induce un effetto ipotermico nei conigli normali e ciò si verifica anche dopo distruzione del centri del caldo. Dosi più elevate riducono l’ipertermia prodotta dalla cocaina e dalla malaria [Jamanchi (187)].
Azione anestetica – Mercier e Delphaut (188) hanno studiato comparativamente, l’azione dell’adrenalina e della yoimbina somministrate per via sottoccipitale e lombare. I risultati ottenuti dimostrano che l’adrenalina a dosi deboli (1-5 goc. di soluz. di cloridrato all’1 %) per via sottoccipitale, non produce alcuna modificazione apprezzabile sulla pressione arteriosa, sul volume del rene o sulla motilità dell’intestino in situ.
Dosi più elevate (2 cc della stessa soluz.) somministrate ad animali con vago sezionato al collo, producono un’ipertensione la cui intensità non è costante in tutti gli animali, ma varia secondo fattori individuali. La pressione può elevarsi lentamente e progressivamente o più rapidamente e raggiungere livelli più elevati. Spesso si nota una vasocostrizione renale e un arresto della motilità dell’intestino in situ. La frequenza respiratoria aumenta subito dopo l’iniezione di adrenalina, ma a volte, questo aumento è preceduto da una fase di rallentamento.
Somministrata per via lombare, l’adrenalina alla dose di 1 cc della stessa soluzione, produce un considerevole effetto ipotensivo, aumento della frequenza respiratoria, leggera costrizione renale, arresto della motilità intestinale e anche fatti convulsivi e spasmi muscolari la cui comparsa in queste condizioni era già stata segnalata dal Bass fin dal 1914 (189). La yoimbina per via sottoccipitale (1-2 mg di cloridrato in 1-2 cc di soluz.) o lombare (1-2 cgr in 1 cc di soluz.), determina regolare caduta della pressione e diminuzione dell’attività respiratoria. Le variazioni del volume del rene seguono passivamente quelle della pressione e l’attività motoria dell’intestino non viene influenzata. Allorché la yoimbina viene somministrata per via lombare, si stabilisce una vera anestesia, che conferma le note proprietà anestetiche di questo alcaloide.
Terapia – La yoimbina ha scarse applicazioni terapeutiche; tuttavia, in epoca relativamente recente, essa è stata impiegata con successo, nella atonia intestinale postoperatoria e in alcuni casi di distonie neurovegetative.
Dieckmann (190) afferma che all’impiego del cloridrato di yoimbina, può essere assegnato un posto degno di rilievo nel trattamento delle sequele operatorie addominali.
La dose di 10 mg di cloridrato di yoimbina somministrata per via endovenosa agli operati addominali, 24 h dopo l’intervento, seguita da una seconda dose alla distanza di 12 h dalla prima, determina, in generale, entro i primi 15 minuti che seguono all’iniezione, una risposta che si manifesta almeno, con l’emissione di gas.
L’innocuità del prodotto, la sua elasticità d’effetti e la lieve reazione, dolorosa che essa produce, sono vantaggi, secondo l’A., per i quali essa dovrebbe essere preferita alla prostigmina.
Analogamente, Deshonnets e Herbaux (191), in base ai risultati ottenuti con l’impiego della yoimbina in 10 casi di ileo paralitico postoperatorio, insistono sull’estrema facilità del trattamento e della sua utilità, dopo che altri mezzi terapeutici hanno fallito.
Un risultato negativo deve far pensare, secondo gli AA. alla presenza di un ostacolo anormale (peritonite, aderenze), per il quale è indispensabile l’intervento chirurgico.
Alle stesse conclusioni è giunto più recentemente Kaiser (192), il quale avendo impiegato la yoimbina in 12 casi di atonia intestinale, ha trovato che essa ha dimostrato un’attività superiore agli altri mezzi terapeutici sino a ora impiegati.
Anche più recentemente Dennemark (193) ha invece sperimentato un preparato in compresse contenenti 2,5 mg di yoimbina, in varie forme di distonie neurovegetative, ottenendo ottimi risultati specialmente in quei casi nei quali è particolarmente interessato il sistema adrenergico. Egli ritiene pertanto, che il suddetto preparato dovrebbe essere in ogni caso, preferito a quelli contenenti gli alcaloidi della Segale cornuta.
La yoimbina è stata preconizzata anche nella terapia dell’ipertensione arteriosa, ma i risultati ottenuti non sono stati convincenti. Secondo Laurence (194), essa può produrre inizialmente un aumento, con sintomi pericolosi.
Tossicità – Dosi elevate di yoimbina causano eccitamento psichico, congestione cerebrale e disturbi gastrici (194). Nei conigli compaiono anche lesioni gravi dell’epitelio renale [Huebner (195)].
L’azione di dosi tossiche si manifesta con gravi e complessi effetti sul miocardio [Muller (196), Tait (197)] con stimolazione generale e successiva paralisi, che interessa particolarmente i centri spinali.
La morte avviene per paralisi respiratoria.
Estratti e preparati vari
Estratto fluido (g 1 = LII gtt).
Dosi: g 0,3-3 pro die.
Preparazioni usuali e formule galeniche
Tintura
Estratto fluido yohimbe………………………………………………. g 20
Alcool di 60°………………………………………………………………. g 80
(g 1,5-15 pro die).
Elisir afrodisiaco
Estratto fluido muira puama
Estratto fluido damiana
Estratto fluido echinacea
Estratto fluido yohimbe…………………………………….. ana g 20
Glicerina e alcool………………………………………………. ana g 120
Tintura vaniglia……………………………………………………….. g 30
Sciroppo arancio………………………………………………………….. g 250
Elisir Garus……………………………………………………….. g 600
(un bicchierino dopo i pasti).
BIBLIOGRAFIA
(1) HESSE O., Ber.. 13. 2308, 1880; Lieb. Ann., ZÌI, 249, 1882 – (2) SPIEGEL L., Chemiker-Ztg., 20, 970, 1896 – C3) PARIS R. e GOUTAREL R„ Ann. pharm trans., Ì6, 15, 1958 – (4) SCHMUTZ J. e LEHNER H„ Helv. Cblm. Asia, 42. 874, 1959 – (5) HERZOG, Ber. disch. pharm. Ges., 15, 4, 1905 – (6) TIHON, Bull. Agrls. Congo Belge, 43, 797, 1952 – (7) RAYMOND-HAMET, J. pharm. shim., 19, 209, 1934 – C8) lANOT M M„ LE MEN I, e HAMMOUDA Y„ Ann. pharm. trans., 14, 341, 1956 – (9) DUPOUY e BEILLE, BuU. sei. pharm.. IO, 201, 1905 – (IO) HAACK E., POPELAK A., SPINGLER H. e KAISER F., Nallirwiss., 41, 479, 1954 – (D) CHAUDHURY e CHAtTERIEE A„ J. Ssi. Ind. Res., B IS. 130, 1959 – (12) ISHIDATE M., OKADA M. e SAITO K., Pharm. Bull., 3, 319, 1955 – (13) HOCHSTEIN F, A., MURAI K. e BOEGEMANN W. H„ J. Ani. Chem. Sos., 77, 3551, 1955 – (14) MUELLER J. M., Experienlia. 13. 479, 1957 – (15) BADER F, E., DICKEL D. F. e SCHLITTLER E., J. Am. Chem. Sos., 76, 1695, 1954 – (16) HOFMANN A., Beh-, Chini. Asia, 37, 849, 1954 – (17) HAACK E, POPELAK A. e SPINGLER H„ Nalurwiss., 43, 328, 1956 . (18) SPIEGEL L„ Ber., 48, 2077, 1915 – (19) SPIEGEL L., ibid., 48. 2084, 1915 – (20) FILIPPI E., Arsh. tarmasol. sper., 23, 107, 129, 1917 – (21) FOURNEAU E, e PAGE H. J., Bull. ssi. pharm., 21. 7, 1914 – (22) EWINS A. I., J. Chem. Sos., lOS, 2739, 1914 – (23) RAYMOND-HAMET, C. R. Sos Biol., 96, 2, 1927 – (24) RAYMOND-HAMET, Compì, rend., 187, 142, 1928 – (25) FOURNEAU E. c FIORE, BuU. Sos. Chini. Franse, 9, 1037, 1911 – (26) FIELD E., J. Chem. Sos., 123, 3003, 1923 – (27) BARGER G. e FIELD E., ibid., 107, 1025, 1915 – C28) WINTERSTEIN E. e WALTER M., Helv. Chim. Asta, 10, 577, 1927 – (29) WARNAT K., Ber., 60, 1118, 1927 – (30) WITKOP
B. e GRASER G., Ueh. Ann., 556, 103, 1944 – (31) BARGER G, e SCHOLZ C, J. Chem. Sos., 614, 1933 – (32) RAYMOND-HAMET, Compì, rend., 212. 305,1941 – (33) SPAETHE. e BRETSCHNEIDER H„ Ber., 63, 2997, 1930 – (34) HAHN G. e JUST F„ ibid., 65, 717, 1932 – (35) MENDLIK F. e WIBAUT J, P„ Res. Trae. Chini., 48, 191, 1929 – (36) MENDLIK F. e WIBAUT I. P„ ibid.. 50, 91, 1931 – (37) MENDLIK F., Pharm. Weekbi., 68, 257, 1931 – (38) BARGER G, c SCHOLZ C„ Heiv. Chim. Asia, 16. 1343, 1933 – (39) WIBAUT I. P. e GASTEL VAN A, 3. P„ Ree. Trav. Chim., 54, 85, 1935 – (40) SCHOLZ C, Helv. Chim. Acta. 18. 923, 1935 – (41) PRUCKNER F. e WITKOP B., Lieb. Ann.. 554, 127, 1943 (42) CLEMO G. R. e SWAN G. A., J, Chem. Soc„ 617, 1946 – (43) JULIAN P. L„ KARPEL W. MAGNANI A, e MEYER E. W., J. Am. Chem. Soc.. 70, 180, 1948 – (44) JULIAN P. L., MAGNANI A., PIKL J. e KARPEL W. J., ibtd.. 70, 174, 1948 – (45) HAHN G., KAPPES E, e LUDEWIG H., Ber., 67; 686, 1934 – (46) MAJIMA R e MURAHASHI S, Proc. !mp. Acad. (Tokio). 10, n. 6, 1934; Colleetei Papers Facuity ScL Osaka ìmp. Vniv., Z 341, 1935; Chem. Abs., 30. 3437, 1936 • (47) SCHWARZ Fxperientia. 6. 330, 1950 – (48) HAHN G. e WERNER H., tieb. Ann., 520, 123. 1935 – (49) HARVBV
D. G., MILLER E. J. e ROBSON W., J. Chem. Soc., 153, 1941 – (50) DEWAR M. J. S, e KING F, bA Nature, 148, 25, 1941 – (51) RAYMOND-HAMET, Compì rend.. 226, 1379, 1948 – (52) WOODWARtf) R B. e WITKOP B., J. Am. Chem. 5oc, 70, 2409, 1948 – (53) SCHLITTLER E. e SPEITEL R, H«Jv^ Chim. Acta. 31, 1199, 194 8 – (54) JULIAN P. L., KARPEL W, J., MAGNANI A, e MEYER E.
J. Am. Chem. Soc., 70, 2834, 1948 – (55) CLEMO G. R c SWAN G. A., Nature. 162, 693, 1948; J. Ch«m; Soc., 487, 1949 • (56) RAYMOND-HAMET, Compì, rend., 221. 387, 1945 – (57) HAHN G, c HANSEL A.. ^ Ber., 71, 2192, 1938 – (58) HAHN G. e HANSEL A., ìbid., 71, 2163, 1938 • (59) FOURNEAU E„ Compì, rend. 148, 1770, 1909; Bull. sci. pharm.. 17, 190, 1910 – (60) JORIO, Ann. Chim. farm., 50, 1939 – (61) HOFMANN A-, Hetv. Chim. Acta. 37, 314, 1954 – (62) JANOT M„ GOUTAREL R., LE HIR A.. AMIN M. e PRELOG V„ Bull. soc. chim. frane.. 1085, 1952 – (63) COOKSON R. C., Chem. Ind., 337, 1953 – (64) JULIAN P. L. e PRINTY H. C., J. Am. Chem. Soc., 71, 3206, 1949 – (65) HAHN G. c BRANDEN- ‘ BERG W., Ber.. 59, 2189, 1926 – (66) HAHN G. e BRANDENBERG W.. ibid., 60, 707, 1927 – (67) STOLL A, e HOFMANN A., J. Am. Chem. Soc., 77, 820, 1955 – (68) DJERASSI C. FISHMAN J., GORMAN M., KUTNEY J, P. e PAKRASHI S. C. ibid., 79, 1217, 1957; Chem. Abs.. 51, 8118, 1957 (69) JANOT M. M., GOUTAREL R. e AMIN M., Compt. rend.. 231. 582, 1950 – (70) RAYMOND- HAMET, ibid., 234, 896, 1952 – (71) LE HIR A., JANOT M M. e GOUTAREL R„ Bull. soc. chimJ Franc., 1027, 1953 – (72) WARNAT K., Ber.. 59, 2388, 1926 – (73) HAHN G. e BRANDENBERG W., ibid., 60, 669, 1927 – (74) HUEBNER C. F. e DICKEL D. F., Sxperientia. 12. 250, 1956 – (75) SPIEGEL L.. Ber., 59, 2706, 1926 – (76) LILLIG, Merk^s Jahrsb.. 42, 20, 1928 – (77) MOOKHERJEE A., J. Indian Chem. Soc.. 18, 33, 1941 – (78) CHATPERJEE A. e PAKRASHI S. C., Naturwiss.. 41. 215, 1954 – (79) MOOKHERJEE A-, J. Indian Chem. Soc.. 20, 11, 1943 – (80) MOOKHERJEE A., ibid.. 23, 6, 1946 – (81) MOOKHERJEE A., ibid.. 28, 29, 1951 – (82) CHATPERJEE A. e PAKRASHI S. C., Sci. and Culture (Calcutta). 18, 443, 1953 – (83) CHATTERJEE A. e PAKRASHI S. C., ibid.. 19. 109, 1953 – (84) CHATPERJEE A. e PAKRASHI S. C., J. Indian Chem. Soc.. 31, 25, 1954 – (85) CHATTERJEE A^ Indiati J. Pharm., 18, 232, 1956 • (86) SVOBODA G. H., J.A.Ph.A., 46, 508, 1957 – (87) JANOT M. M. e GOUTAREL R., Compt. rend.. 220, 617, 1945 – (88) URIBE VERGARA B., J. Am Chem. Soc., 77, 1864, 1955 – (89) CHATrERJEE A. e BOSE S., Experientia. 10. 246, 1954 – (90) CHATTERJEE A. c BOSE S-, Sci. and Culture (Calcutta). 19, 512, 1954 – (91) GHOSH B, P., Naturwiss.. 45. 365, 1959; Chem. Abs.. 53, 4655 d, 1959 – (92) HOCHSTEIN F. A., J. Org. Chem., 21. 1516, 1956 – (93) LE HIR A. c GOUTAREL R., Bull. soc. chim. Frane-, 1023, 1953 • (94) CHAITERJEE A., BOSE A, K. e PAK- RASHI S-, Chem. Ind.. 491, 1954 – (95) HEINEMANN M., Ber.. 67, 15, 1934 – (96) HOFMANN A./ Helv. Chim. Acta. 38, 536, 1955 – (97) KIMOTO S. c INOUE H., J. Pharm. Soc. Japan. 62. 95, 1942 – (98) KIMOTO S. c OKAMOTO M., Pharm. Bull.. 3. 392, 1955 – (99) SIDDIQUI S. e SIDDIQUI R. H., J. Indian Chem. Soc.. 8, 667, 1931; 9, 539, 1932; 12. 37, 1935 – (100) NEUSS N., BOAZ H. E, e FORBES J. W„ J. Am. Chem. Soc.. 76, 3234, 1954 – (101) JANOT M, M. c LE MEN J„ Compt. rend., 239, 1311, 1954 – (102) GOUTAREL R. e LE HIR A., Buil soc. chim. Frane., 909, 1951 – (103) WENKERT E. e ROYCHAUDHURI D. K., J. Am. Chem. Soc.. 79, 1519, 1957 – (104) WENKERT E. e ROYCHAUDHURI D. K., ibid., 80. 1613, 1958 – (105) NEUSS N. c BOAZ H. E., J. Org. Chem. 22, 1001, 1957 – (106> TAMELEN VAN E. E. c PLACEWAY C., J. Am. Chem. Soc.. 83, 2594, 1961 – (107) RAYMOND- HAMET, Compt. rend.. 197. 860, 1933 – (108) KARRER P. c SALOMON H^ Helv. Chim Acta. 9, 1059, 1926 – (109) KARRER P. e ENSLIN P., ibid.. 32, 1390, 1949 – (110) CHATTERJEE A. c KARRER P-, ibid.. 33, 802, 1950 – (111) PRELOG V., JANOT M. M. c GOUTAREL R., Compt. rend.. 232, 1305, 1951 – (112) JANOT M. M„ GOUTAREL R. c PRELOG V., Helv. Chim. Acta. 34, 1207, 1951 – (113) JANOT M. M. c GOUTAREL R., Bull. soc. chim. Frane.. 588, 1951 – (114) KARRER P., SCHWYZER R. c FLAM A.. Helv. Chim. Acta, 35. 851, 1952 – (115) KARRER P. c ST. MAINONI, ibid.. 36. 127, 1953 – (116) ANDERSON R. M., CLEMO G. R. c SWAN G. A., J. Chem. Soc.. 2962, 1954 – (117)
LEE T. B. c SWAN G. A., ibid.. 771, 1956 – (118) JANOT M M., GOUTAREL R., LE HIR A.,
TSATSAS G, c PRELOG V., Helv. Chim. Acta, 38, 1073, 1955 – (119) WENKERT E. c ROYCHAU- DHURI D. K, J. Am. Chem Soc., 78. 6417, J956 – (120) TAMELEN VAN E. E., ALDRICH P. E. e KATZT. J„ Chem. Ind., 793, 1956 – (121) TAMELEN VAN E. E. c HESTER J. B., J. Am. Chem. Soc.. 81. 3805, 1959 – (122) BRANDT W„ Arch. Pharm., 260. 49. 1922 – (123) PERROT E. c RAYMOND-
HAMET, BuU. sci. pharm. 39, 593, 1932 – (124) FELDHOFF, Pharm. Ztg.. 70. 864, 1925 – (125)
SCHOMER, Pharm. Zentralh., 63. 385, 1922 – (126) HAHMANN. Apoth. Ztg., 42, 1385, 1927 – (127) PARIS R. R c LETOUZEY R, J. agr. trop. botan. appi.. 7. 256, 1960 – (128) PARIS R. R e LETOUZEY R., ibid.. 4. 31, 1957; Chem. Abs.. $4. 13275, 1960 • (129) OSSERVATE e LOEWY, cit. in EHspensatory of The U.StA., Uppìocot Comp. Ed., PhìUdelphia, London, Montreal 1955, p. 1932 – (130) MUELLER’, Arch. Interri. Pharmacodyn. Thfr.. 65, 1907 • (131) RAYMOND-HAMET, Arch. exp, Path. u. Pharmakoi. 184, 680, 1937 – (132) FUCO N. W. e CROSS, Endocrinotogy. 31, 529, 1942 – (133) RUMRY, cit. da SOLLMANN T. in A Manual o( Pharmacology and it$ application lo Therapeutics and Toxicology 7> ed. W. B. Saunders Comp., Philadelphia London 1950, p. 278 – (134) RAYMOND-HAMET, Compt. rend.. 180, 2074, 1925 – (135) FILIPPI R, Arch. It. Farmacoì. Sperim.. 23, 107, 1917 ^ (136) RAYMOND- HAMET, Buil. Acad. Med.. 91, 145, 1924 – (137) RAYMOND-HAMET. C.R. Soc. BÌol., 96, 2, 1927 – (138) RAYMOND-HAMET. Compt. rend.. 182. 170, 1925 – (139) RAYMOND-HAMET, ibid.. 183, 485, 1926 – (140) RAYMOND-HAMET, ibid., 183, 1124, 1926 – (141) RAYMOND-HAMET. ibid., 184, 1672, 1927 – (142) RAYMOND-HAMET, ibid.. 190, 9TÌ4, 1930 – (143) RAYMOND-HAMET, C. R. Soc. Biot.. 108, 963, 1931 – (144) RAYMOND-HAMET, Comunicai. all’Acad. de Sci. 18 febbr. 1952 in Presse méd., 60, 304, 1952 – (145) HUTCHINSON e coU., dt. da SOLLMANN l.c., vedi (133) – (146) BARRY D. T. e CHAUCHARD. C. R. Soc. Bioi., 125, 217, 1937 – (147) BARRY D. T., Arch. Int. Pharmacodyn. et Thér.. 55, 385, 1937 – (148) KOPPANYI T., LINECAR C. R. e HERWICK R. P., Fed. Proc,, 2, 83, 1943 – (149) JONESCU, Wiener Kìin, Woch.. 25, M3, 1908 – (150) DALE, J. Physioi., 34, 176, 1906 – (151) DALE, ibid., 46, 293, 1913 – (152) HOUSSAY R. D. e MOLINELLI F. A., C. R. Soc. Biot., 95, 808, 1926 – (153) GLEY R e CZARNECKI EDW, ibid„ 97, 1156, 1927 – (154) LOEPER M. e LEMAIRE A., ibid., 105. 109, 1930 – (155) LOEPER M., VILLARET M., JUSTIN-BESANCON L., LEMAIRE A. e CACHERÀ R., ibid.. 104, 1203, 1930 – (156) HEYMANS L. e REICNERS P., ibid., 101, 257, 1929 – (157) NICKERSON M., Pharmacoi. Rev„ 1, 27, 1949 – (158) HEYMANS C. e BOUCKAERT J. J., C.R. Soc. Bioi., 103, 923, 1930 • (159) BOUCKAERT J. J. e CZARNECKI EDW., Ann. Soc. Scient, Bruxelles 42* serie. 91, 1947 • (160) RABE, cit. da SOLLMANN, vedi (133) – (161) MEYER F.. idem. – (162) WEGER P., Rev. Pharmacoi. Ther. exp.. 1, 173, 1929, cit. da SOLLMANN, idem – (163) GERSHON S. e LANG W. I., Arch. Int. Pharmacodyn. et Thér.. 135, 31, 1962 – (164) YONKMANN F. F., STILWELL D, e JEREMIAS R., J. Pharmacoi. exp. Ther.. 81, 111, 1944 – (165) HUTCHINSON M., SHOW F. H e WRAGGE W.. Aust. J, exp. Bioi. e med. Sci.. 20, 69, 1942 – (166) DELGA J. e HAZARD R., Arch. ìnt. Pharmacodyn et Thér., 117, 88, 1958 – (167) HOLMBERG G. e GERSHON S., dt. da GERSHON e LONG in (163) – (168) SHAW E. e WOLLEY D. W., J. Bioi. Chem., 203. 279, 1953 – (169) NORTHO VER B. J. e SUBRAMANIAN G.; Brit. J. Pharmacoi.. 18, 346, 1955 – (170) SIGG E. B., Can. Psychlat. Assoc. J., 4. 575, 1959 – (171) HOLMBERG G., GERSHON S. e BECK L. H., Nature, 193, 1313, 1962 – (172) BARIETY M. e KOHLER D., C. R. Soc. Bioi. 130, 1009, 1939 – (173) LOEWY A. e ROSENBERG, cit. da SOLLMANN, vedi (133) – (174) RAYMOND-HAMET, Compt. rend., 185, 610, 1927 – (175) CHAUCHARD A. B. e CHAUCHARD P.. C R. Soc. Bioi. 126, 1171, 1937 – (176) DELGA J. e HAZARD R, ibid.. 148, 1802, 1954 – (177) DELGA J. e HAZARD R., ibid,, 149, 1756, 1955 – (178) CORMIER R., VAN DEN DRIESSCHE J., TREBAUL L. e LE BARS R., Thérapie. 15. 247, 1960 – (179) FENN W. O, e ASANO T., Am. J. Physioi. 185, 567, 1956 – (180) D’SILVA J. L., J. Physioi (London). 86, 219, 1936 • (181) NITZESCU J. J., C. R. Soc. Bioi. 98. 1482, 1928 – (182) HANSON S., ibid.. 101, 601, 1929 – (183) HANSON S., ibid.. 101, 603, 1929 – (184) HANSON S.. ibid.. 101, 605, 1929 – (185) THOUVENAT J. e HARICHAUX P., ibid., 155. 1986, 1961 – (186) FUGO N. W., Endocrinotogy, 34, 143, 1944 – (187) JAMANCHI, 1928, cit. da SOLLMANN, vedi (133) – (188) MERCIER F, e DELPHAUT J., C.R. Soc. Bioi. 123. 24, 1936 – (189) RASS, Zentr. Biochem. Bioph., 17, 639, 1914 – (190) DIECKMANN D. G., Soc. Med. Chir. Des. Hópii. tibres. 3, VI, 1950; Presse Méd.. 58, 859, 1950 – (191) DESHONNETS G. e HERBAUX J., Soc. Med. Anat. din. de Lille, 12. VII, 1950, in Presse méd.. 59, 456, 1951 – (192) KAISER R., Med. Weit., 16. 856, 1960 • (193) DENNEMAKER H. G., Ther. der Gegenw.. 3, 82, 1950 – (194) SOLLMANN T., vedi (133) – (195) HUEBNER, cit. da SOLLMANN, vedi (133) – (196) MUELLER F.. Arch. int. Pharmacodyn et Thér.. 65, 1907 – (197) TAIT, cit. da SOLLMANN, vedi (133).